
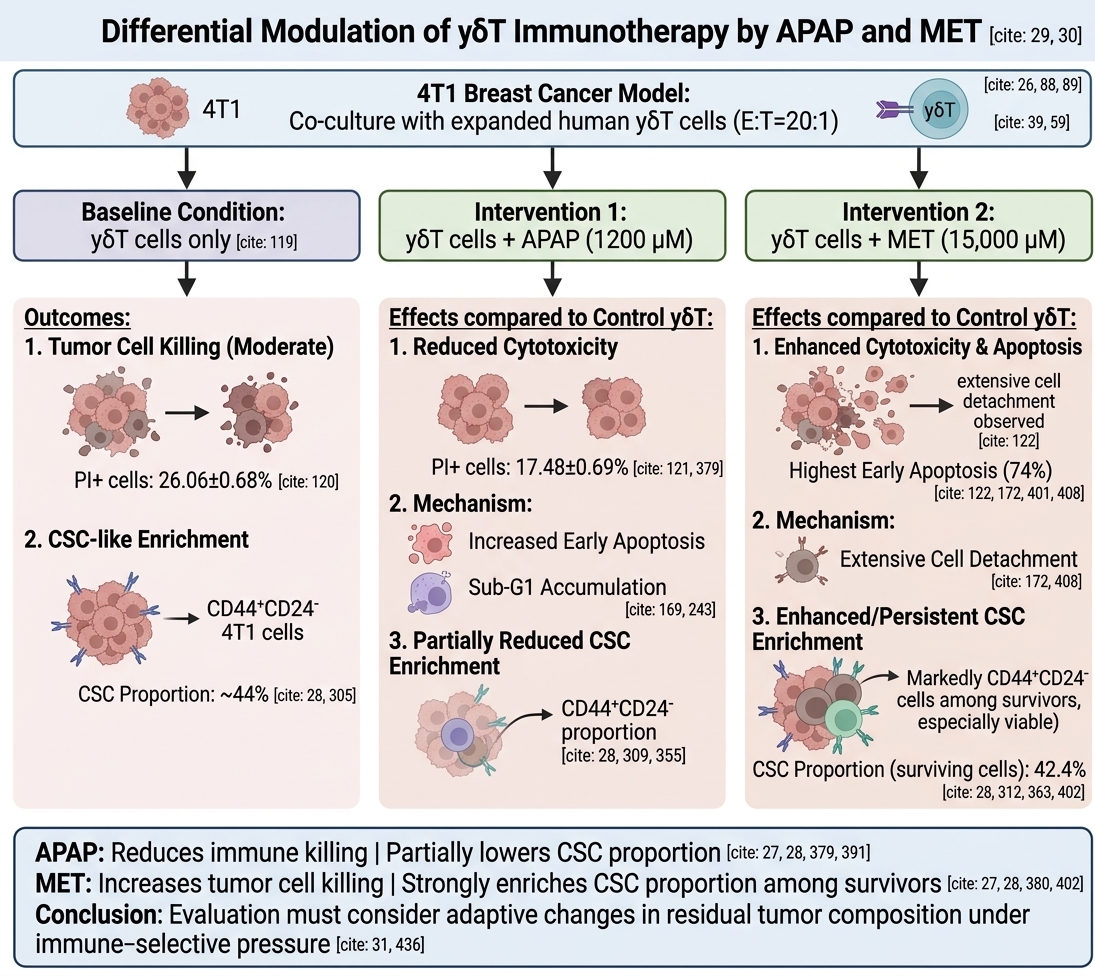

Acetaminophen and Metformin Differentially Modulate γδT Cell-Mediated Antitumor Activity and CSC-like Population Enrichment in the 4T1 Breast Cancer Model
- VNUHCM-US Stem Cell Institute, University of Science, Ho Chi Minh City, Viet Nam
Abstract
Background: γδT cells are emerging as promising effectors in cancer immunotherapy due to their potent MHC-independent cytotoxicity. However, the impact of commonly used pharmacological agents on γδT cell-mediated antitumor activity remains poorly understood. In particular, acetaminophen (APAP), widely used in supportive care, and metformin (MET), a metabolic modulator with anticancer potential, may influence tumor–immune interactions and therapeutic outcomes.
Methods: Using the highly aggressive murine 4T1 breast cancer model, which exhibits TNBC-like characteristics including enhanced metastatic behavior and stemness-associated properties, we investigated the effects of APAP and MET on γδT cell-mediated antitumor activity and tumor-cell adaptation under immune-selective pressure. Co-culture systems (E:T = 20:1) were established, and tumor cell death, apoptosis, and cell cycle distribution were assessed by flow cytometry (PI, Annexin V/PI, PI/RNase). Cancer stem cell (CSC)-like populations (CD44⁺CD24⁻) were analyzed by flow cytometry (FCM), and viability of CSC-like and non-CSC subpopulations were evaluated using the AlamarBlue assay.
Results: γδT cells significantly induced cytotoxicity in 4T1 cells; however, APAP reduced γδT-mediated killing despite promoting early apoptosis and increasing Sub-G1 accumulation. In contrast, MET exhibited strong direct cytotoxicity and markedly enhanced apoptotic induction, particularly in combination with γδT cells. Notably, both APAP and MET were associated with enrichment of CSC-like populations, with MET showing the strongest effect, especially among surviving cells. Functional assays revealed that CD44+CD24− CSC-like populations were not intrinsically resistant to γδT-mediated killing, but treatment-induced selective pressure reshaped tumor cell composition.
Conclusion: APAP and MET differentially modulated γδT cell antitumor activity in breast cancer. While APAP was associated with reduced immune-mediated tumor killing, MET enhanced tumor cell killing but simultaneously promoted CSC-like population enrichment among surviving cells. These findings suggest that therapeutic evaluation should consider not only immediate tumor cell killing but also adaptive changes in residual tumor-cell composition under immune-selective pressure.
Introduction
Breast cancer remains the most frequently diagnosed malignancy and one of the leading causes of cancer-related mortality among women worldwide. Despite substantial advances in surgery, chemotherapy, endocrine therapy, targeted therapy, and immune checkpoint blockade, therapeutic resistance, tumor recurrence, and metastatic progression continue to limit long-term survival, particularly in aggressive subtypes such as triple-negative breast cancer (TNBC)1,2,3. A major contributor to these clinical challenges is the persistence of cancer stem cell (CSC)-like populations, which possess enhanced self-renewal capacity, resistance to conventional therapies, metastatic potential, and immune escape properties4,5,6. In breast cancer, CSC-like populations are frequently associated with the CD44CD24 phenotype and are strongly associated with poor prognosis and treatment failure7,8,9,10.
Among emerging immunotherapeutic strategies, γδT cells have attracted increasing attention due to their potent major histocompatibility complex (MHC)-independent antitumor activity. Unlike conventional αβ T cells, γδT cells recognize stress-induced ligands, phosphoantigens, and transformed cells without requiring classical antigen presentation, allowing rapid and broad immune surveillance against tumor cells. Their cytotoxic effects are mediated through perforin/granzyme release, Fas/FasL interactions, TRAIL signaling, and secretion of pro-inflammatory cytokines such as IFN-γ and TNF-α11,12,13. Recent studies have highlighted the therapeutic relevance of γδT cells in solid tumors, particularly TNBC, where they can target both differentiated tumor cells and CSC-like populations4,11,12. However, γδT cell function is highly influenced by the tumor microenvironment, and both antitumor and protumor roles have been described depending on local immune context and metabolic conditions12,14,15,16.
Metabolic modulation has emerged as an important determinant of tumor–immune interactions. Metformin (MET), a widely prescribed anti-diabetic drug, has gained significant interest as a potential anticancer agent due to its ability to inhibit mitochondrial complex I, disrupt oxidative phosphorylation, activate AMPK signaling, and suppress anabolic pathways required for tumor growth. In breast cancer, MET has been reported to reduce proliferation, induce apoptosis, and modulate CSC maintenance, although its effects on residual stem-like tumor populations remain controversial17,18,19,20. Importantly, MET may also reshape immune responses by altering both tumor cell metabolism and immune effector cell function, suggesting potential implications for combination immunotherapy19,20.
In contrast, acetaminophen (APAP, paracetamol), one of the most commonly used analgesic and antipyretic agents in cancer supportive care, has received far less attention regarding its immunological consequences. APAP is widely administered to manage pain and fever during cancer treatment, yet increasing evidence suggests that it may influence redox balance, mitochondrial function, and immune regulation beyond its conventional symptomatic use21,22,23. APAP-induced glutathione depletion and mitochondrial oxidative stress can promote apoptotic signaling, but excessive oxidative stress may also interfere with immune synapse formation, death receptor signaling, and cytotoxic lymphocyte function24,25. Given the routine use of APAP in oncology settings, its potential impact on antitumor immune responses deserves closer investigation.
In this study, we investigated the effects of APAP and MET on γδT cell-mediated antitumor activity using the highly aggressive murine 4T1 breast cancer model. Compared with luminal breast cancer models, 4T1 cells exhibit enhanced metastatic potential, increased stemness-associated properties, and distinct responses to therapeutic stress. Therefore, this study aimed not only to evaluate how APAP and MET influence γδT-mediated cytotoxicity, but also to examine tumor-cell adaptation under immune-selective pressure, including differential susceptibility of CSC-like and non-CSC subpopulations, apoptotic progression, cell cycle redistribution, and CSC-like population enrichment. Our findings provide additional insights into how commonly used pharmacological agents may reshape antitumor immune responses, influence residual tumor-cell composition, and affect CSC-associated adaptive responses in an aggressive TNBC-like setting.
Materials and Methods
Cell culture and reagents
The breast cancer cell line 4T1 was maintained in RPMI 1640 medium (Gibco, Cat# 61870-036) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco, Cat# A5256701) and 1X antibiotic-antimycotic solution (Gibco, Cat# 15240-062). Cultures were incubated at 37°C in a humidified 5% CO atmosphere, with medium replenished every 48 h. Subculturing was performed utilizing TrypLE Express (Gibco, Cat# 12604021) upon reaching 80-90% confluence. Routine testing was performed to ensure cell lines were free of Mycoplasma contamination prior to functional assays. APAP (Sigma-Aldrich, Cat# A7085) was dissolved in DMSO to generate a 5 M stock, ensuring the final DMSO concentration in all experimental conditions strictly remained ≤ 0.1% (v/v) to circumvent vehicle-induced toxicity. MET (Sigma-Aldrich, Cat# 317240) was prepared as a 1 M stock in sterile nuclease-free water.
Ex Vivo expansion and phenotypic validation of primary γδT cells
Isolation and expansion
Peripheral venous blood samples (15 mL) were obtained from healthy adult volunteers. All procedures involving human participants were performed in accordance with the Declaration of Helsinki and were approved by the Institutional Ethics Committee. Written informed consent was secured from each donor before sample collection.
Peripheral blood mononuclear cells (PBMCs) were separated using density-gradient centrifugation (1500 × g for 15 min). To induce selective activation of γδT cells, PBMCs were first maintained in GDTCult D1-3 medium (Regenmedlab, Cat# 354) for 48 h. The suspension cell fraction was then collected and transferred into gas-permeable culture bags for further expansion in GDTCult D3-21 medium, with fresh medium supplementation every 3 days. After 21 days of culture, the expanded γδT cells were collected and used for subsequent immunological analyses.
Flow cytometric quality control
Before performing functional co-culture experiments, the purity of the expanded effector cell population was carefully assessed. A total of 0.5–2 × 10 cells were stained with PerCP-Cy5.5-conjugated Mouse Anti-Human TCR γδ antibody (BD Pharmingen, Cat# 564157). Flow cytometric analysis was carried out using a BD FACS Melody system (BD Biosciences) to determine the proportion of TCR γδ+ cells. Expanded cell products containing at least 50% TCR γδ+ cells were considered sufficiently qualified and were selected for subsequent functional assays (Supplementary, Figure S1).
Pharmacological modulation and metabolic stress modeling
To establish in vitro models of acute, targeted metabolic and redox stress, the half-maximal inhibitory concentrations (IC) of APAP and MET on 4T1 cells were predetermined. Cells (2,000 cells/well) were exposed to dose-escalations of APAP (0–1,000 µM) or MET (0–20,000 µM). Viability was quantified at 24, 48, and 72 h via the AlamarBlue assay (Thermo Fisher, Cat# DAL1025). Based on nonlinear regression analyses, stress-inducing concentrations corresponding to their 48-h kinetic profiles (1,200 μM APAP and 15,000 μM MET) were selected for downstream immune co-cultures to evaluate modulatory effects without inducing absolute target cell ablation (The IC₅₀ values were previously determined by our team). These concentrations were selected to generate measurable biological responses and immune-selective pressure within the in vitro co-culture system rather than to directly reproduce physiological plasma exposure levels.
Multiparametric assessment of immune-mediated cytotoxicity and cell cycle dysregulation
Co-culture conditions
4T1 target cells were seeded into T75 culture flasks and allowed to attach for 48 h before co-culture. Expanded primary γδT cells were then added at an effector-to-target (E:T) ratio of 20:1. The co-culture system was maintained for an additional 48 h under either standard culture conditions or in the presence of pharmacological treatments, including 1,200 μM APAP and 15,000 μM MET. Human donor-derived γδT cells were used as effector cells in this exploratory co-culture system to investigate tumor-cell responses under immune-selective conditions.
Viability and apoptotic mapping
To quantify overall target cell death and distinguish apoptotic stages and necrotic cell populations, cells were dual-stained with Annexin V-FITC and Propidium Iodide (PI) using the Annexin V Apoptosis Detection Kit (BD Pharmingen, Cat# 556547).
Cell cycle profiling
Cell cycle distribution and Sub-G1 DNA fragmentation were evaluated using PI/RNase Staining Buffer (BD Pharmingen, Cat# 550825). Briefly, cells were fixed overnight in 70% cold ethanol at −20°C, followed by staining according to the manufacturer’s instructions. Flow cytometric data were acquired and analyzed using FlowJo v10.8 software (BD Biosciences) to quantify the percentages of cells in Sub-G1, G1/G0, S, G2, and >G2 phases.
Following co-culture, the non-adherent fraction containing the majority of γδT effector cells was removed prior to tumor cell collection. For all flow cytometry-based co-culture analyses, single cells were located based on differences in Forward Scatter (FSC) and Side Scatter (SSC) characteristics. Sequential gating strategies were applied to exclude debris and remove doublets using SSC-W vs. SSC-H and FSC-W vs. FSC-H discrimination, ensuring accurate single-cell analysis of the target tumor population.
Isolation and analysis of CSC-like subpopulations
4T1 cells were collected by centrifugation (500 × g for 5 min), washed twice with PBS, and resuspended in sorting buffer consisting of PBS supplemented with 0.1% BSA. Cells were then stained with PE-conjugated anti-mouse CD44 monoclonal antibody (clone IM7; BD Biosciences, Cat# 12-0441-82) and FITC-conjugated anti-mouse CD24 monoclonal antibody (clone M1/69; BD Biosciences, Cat# 11-0242-82), using 5 µL of each antibody per sample, and incubated for 20 min at 4–8°C in the dark.
Following staining, cells were washed twice with sorting buffer (500 × g for 5 min), resuspended in 0.5–1 mL of buffer, and subjected to fluorescence-activated cell sorting (FACS) using a BD FACS Melody system (BD Biosciences). The CD44CD24 fraction was identified and isolated as the CSC-like enriched population. Sorted cells were subsequently resuspended either in complete culture medium containing 2× antibiotic–antimycotic for further in vitro culture or in PBS for downstream experimental analyses.
For cytotoxicity assays, sorted CD44CD24 (CSC-like) and non-CSC (CD44CD24 and CD44CD24) populations were seeded into 96-well plates at a density of 5,000 cells per well and allowed to attach overnight. Effector immune cells were then added at effector-to-target (E:T) ratios of 0:1, 10:1, 20:1, 30:1, 40:1, and 50:1. After 48 h of co-culture, effector cells were removed by gentle PBS washing, and the remaining viability and proliferative capacity of the 4T1 subpopulations were determined using the AlamarBlue assay.
Statistical analysis
All quantitative data are expressed as mean ± standard deviation (SD) from at least three independent biological replicates (n = 3). Statistical analyses were performed using GraphPad Prism version 10.4.1 (GraphPad Software, San Diego, CA, USA).
For comparisons between two groups, an unpaired two-tailed Student’s t-test with Welch’s correction was used. Multiple-group comparisons were analyzed using one-way or two-way analysis of variance (ANOVA), followed by Tukey’s multiple comparison post hoc test with multiplicity-adjusted P values to control the family-wise error rate. A family-wise significance threshold of α = 0.05 was applied. Statistical significance was defined as p < 0.05.
Results
APAP reduced γδT-mediated cytotoxicity in 4T1 cells
The percentage of 4T1 cell death was evaluated by propidium iodide (PI) staining followed by flow cytometry (FCM) analysis under co-culture conditions with γδT cells, in the presence or absence of APAP and MET (Figure 1). Co-culture with γδT significantly increased the proportion of PI-positive 4T1 cells (26.06 ± 0.68%) compared to the control group (4.11 ± 0.33%, p < 0.0001), confirming the cytotoxic activity of γδT.
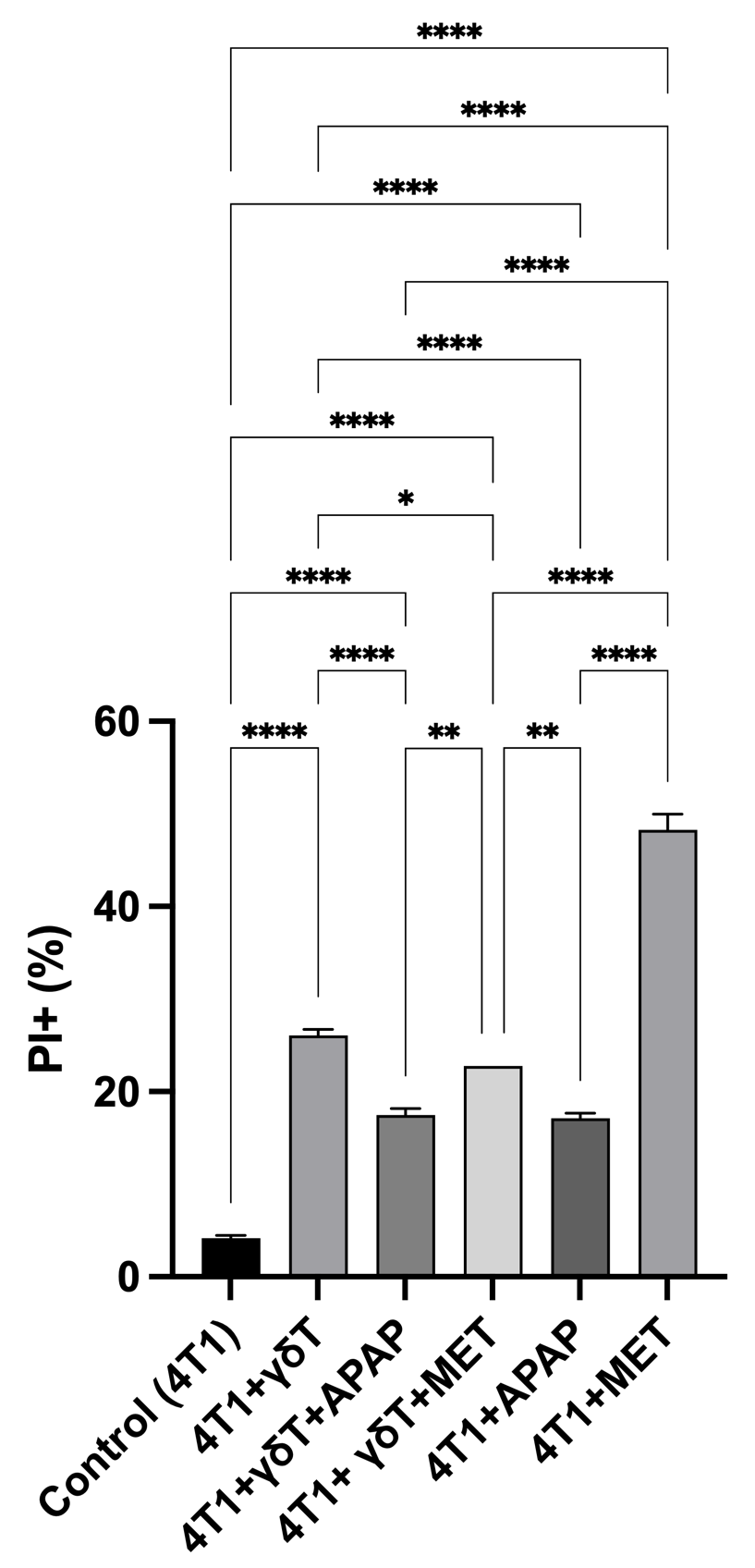
APAP attenuated γδT-mediated cytotoxicity against 4T1 cells. Cell death was assessed by propidium iodide (PI) staining followed by flow cytometry after co-culture with γδT cells. Data are presented as mean ± SD (n = 3). Co-culture with γδT significantly increased the percentage of PI-positive 4T1 cells compared to the control group, confirming the cytotoxic activity of γδT. This effect was reduced in the presence of APAP and partially reduced in the γδT +MET group. APAP alone induced moderate cell death, whereas MET alone showed a markedly higher level of cytotoxicity. The relatively lower PI-positive percentage observed in the γδT +MET group may be associated with extensive cell detachment during co-culture, which could lead to underestimation of total cell death. Statistical significance between groups is indicated as
Upon APAP treatment, the percentage of dead 4T1 cells significantly decreased to 17.48 ± 0.69% (p < 0.0001), suggesting that APAP was associated with reduced tumor killing under γδT co-culture conditions. Similarly, the APAP-only group also exhibited a comparable level of cell death (17.08 ± 0.56%), indicating that APAP alone induced only moderate cytotoxicity toward 4T1 cells.
In contrast, MET alone resulted in a markedly higher proportion of PI-positive cells (48.30 ± 1.66%), suggesting a strong cytotoxic effect under these conditions. Interestingly, the combination of γδT and MET yielded a lower apparent cell death rate (22.78%), comparable to the γδT+APAP group but still below that of MET alone.
It is important to note that PI-based measurements primarily reflect membrane-compromised cells remaining in the analyzed population. As shown in Figure 2, the combination of γδT and MET led to extensive cell detachment, suggesting that the relatively lower PI-positive percentage in this group may underestimate the overall cytotoxic effect due to loss of detached dead cells during sample processing.
Overall, these data suggest an association between APAP treatment and reduced tumor killing under γδT co-culture conditions, whereas MET exhibits strong independent cytotoxicity but may interfere with accurate quantification in co-culture conditions due to increased cell detachment.

The morphology of 4T1 cells following treatment with γδT, with or without APAP and MET. Representative bright-field microscopic images (100x magnification) illustrating the drastic reduction in adherent cellular density following 48-hour exposure to established IC50 drug concentrations, with or without γδT cell co-culture (E:T = 20:1).
Inverted microscopy revealed differences in adherent cell density across treatment conditions. In the control group (4T1 alone), cells nearly covered the entire culture surface by 48 h. Upon adding γδT to the culture, cell density reduced dramatically. Adding APAP did not significantly change the morphology of the cells comparing to the 4T1+ γδT, but adding MET destroyed almost all cancer cells. 4T1 cell’s density in APAP or MET alone treatment was markedly decreased comparing to the control but higher than other treatments. Especially, the cells treated in MET tended to cluster together more tightly (Figure 2).
APAP and MET enhance γδT-induced apoptosis in 4T1 cells
Apoptotic cell death of 4T1 cells was evaluated by flow cytometry using dual staining with Annexin V-FITC and Propidium Iodide (PI) after co-culture with γδT cells, in the presence or absence of APAP and MET (Figure 3). The percentages of early apoptosis, late apoptosis, and necrosis were analyzed to determine the effects of γδT-mediated cytotoxicity and drug treatment on cell death progression. Compared with the control group, co-culture with γδT alone increased both early apoptosis (24.73 ± 0.49% vs. 15.47 ± 3.69%) and late apoptosis (23.00 ± 0.66% vs. 3.51 ± 0.32%), indicating that γδT effectively induced apoptotic cell death in 4T1 cells. Necrosis was also slightly increased (3.06 ± 0.07% vs. 0.60 ± 0.17%), although apoptosis remained the dominant mode of cell death.
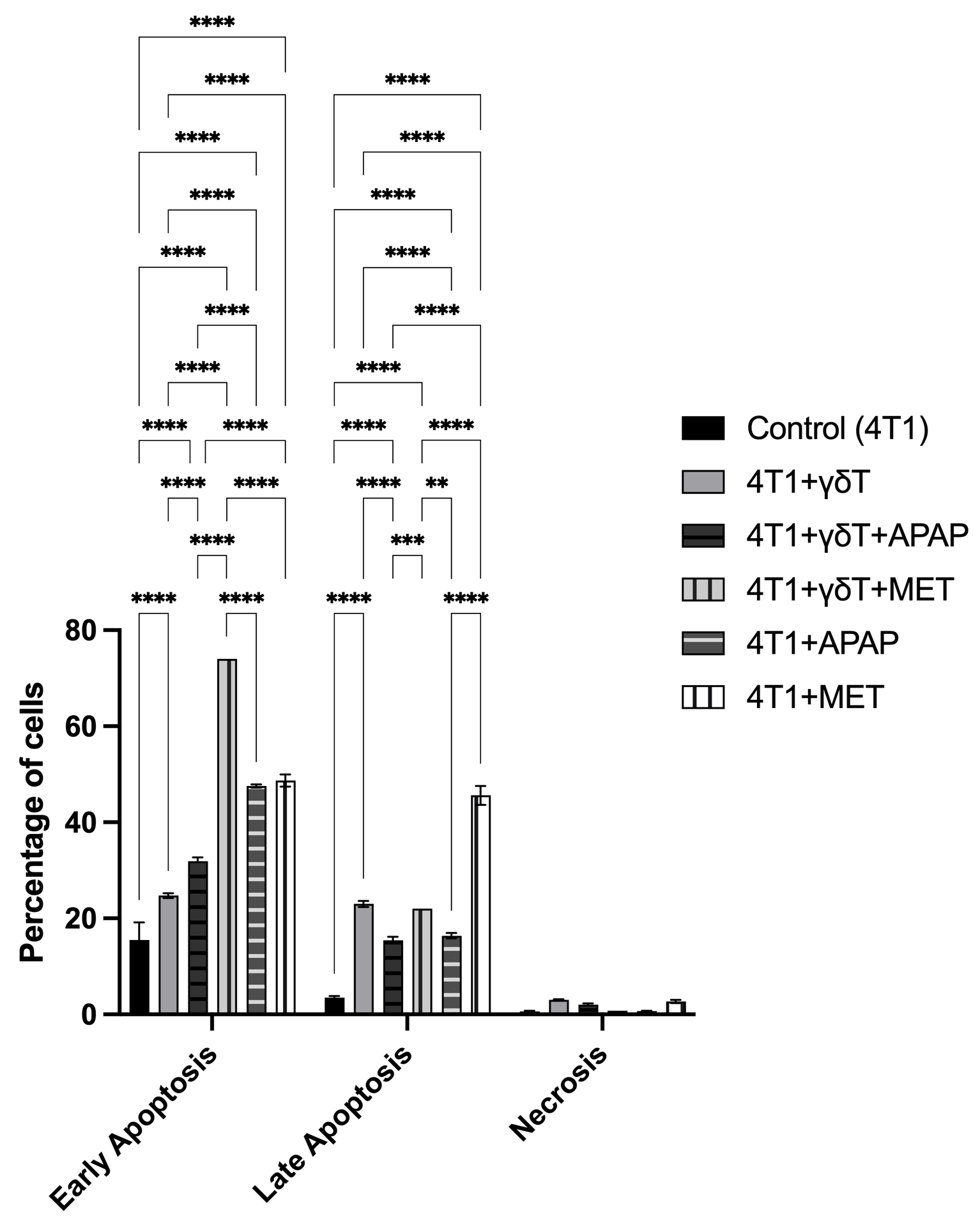
APAP and MET enhance γδT-mediated apoptotic cell death in 4T1 cells. Apoptotic analysis of 4T1 cells after co-culture with γδT cells in the presence or absence of APAP and MET using Annexin V-FITC/PI dual staining. Cell death was evaluated by flow cytometry to determine the percentages of early apoptosis, late apoptosis, and necrosis. Data are presented as mean ± SD (n=3). Co-culture with γδT increased both early and late apoptotic populations compared to the control group. APAP treatment further enhanced early apoptosis, while MET alone and in combination with γδT induced markedly increased apoptotic cell death, particularly in the early apoptotic phase. Necrotic populations remained relatively low across all groups, indicating that apoptosis was the predominant mode of cell death. Statistical significance between groups is indicated as p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
The addition of APAP to the γδT co-culture further increased early apoptosis to 31.90 ± 0.79%, while late apoptosis decreased to 15.43 ± 0.75%, suggesting that APAP may enhance the initiation of apoptosis but reduce the progression to late apoptotic stages. Similarly, APAP alone markedly elevated early apoptosis (47.57 ± 0.31%) with a moderate increase in late apoptosis (16.39 ± 0.57%), confirming its direct pro-apoptotic effect on 4T1 cells.
MET treatment also induced substantial apoptosis. MET alone resulted in high levels of both early apoptosis (48.70 ± 1.27%) and late apoptosis (45.60 ± 1.98%), indicating strong cytotoxic activity. Notably, the combination of γδT and MET showed the highest proportion of early apoptosis (74.00%) while maintaining elevated late apoptosis (22.01%), suggesting that MET may further enhance γδT-mediated apoptotic cell death in 4T1 cells. Necrotic populations remained relatively low across all treatment groups, indicating that apoptosis was the primary mechanism of cell death.
Overall, these results demonstrate that both APAP and MET enhance apoptotic cell death in 4T1 cells, with MET showing a stronger pro-apoptotic effect, particularly when combined with γδT.
Differential susceptibility of 4T1 cell subpopulations to γδT cell-mediated cytotoxicity
Cell viability of total 4T1 cells, 4T1 CSC-like populations (CD44CD24), and 4T1 non-CSCs was evaluated using the AlamarBlue assay following co-culture with γδT cells at different effector-to-target (E:T) ratios (Figure 4). Overall, γδT cells reduced the viability of all three 4T1 populations in an E:T ratio-dependent manner. In the unsorted 4T1 population, cell viability decreased progressively from 100.00 ± 14.49% at baseline to 53.87 ± 6.80% at an E:T ratio of 50:1.
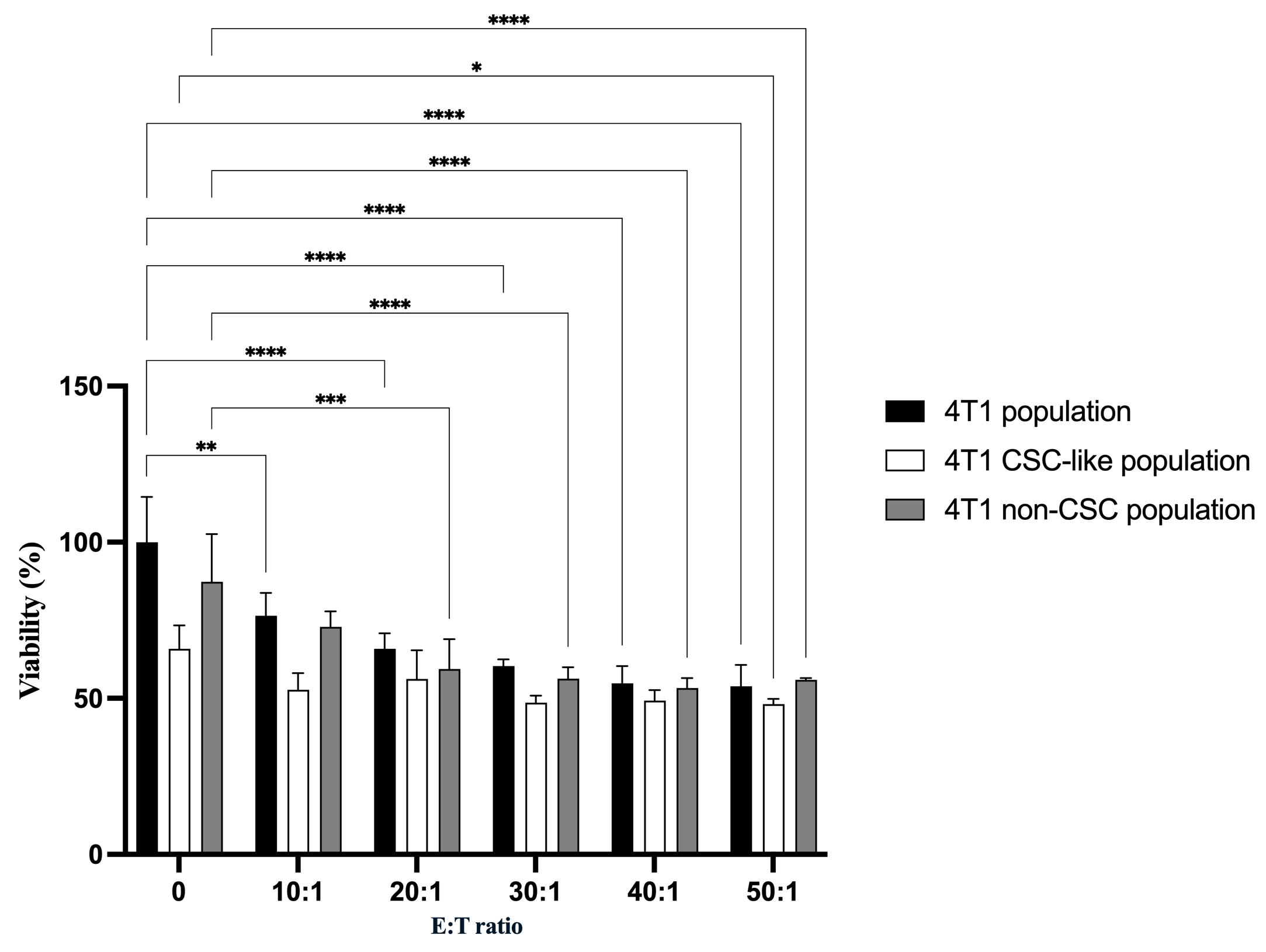
Differential viability of 4T1 cell subpopulations following co-culture with γδT cells. γδT-mediated cytotoxicity against 4T1 tumor cell subpopulations at increasing effector-to-target (E:T) ratios. Cell viability of total 4T1 cells, CD44+CD24− CSC-like populations, and non-CSC populations (isolated from the parental population) was determined by AlamarBlue assay following co-culture with γδT cells. Data are shown as mean ± SD (n = 3). A dose-dependent reduction in viability was observed across all cell populations. Statistical comparisons between indicated groups are denoted by p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
When comparing subpopulations, 4T1 CSC-like populations consistently exhibited lower viability than the total 4T1 population across most E:T ratios, indicating a higher susceptibility to γδT-mediated cytotoxicity. In contrast, 4T1 non-CSCs showed relatively higher viability than CSC-like populations, particularly at lower E:T ratios. Specifically, despite the lower viability, CD44CD24 populations were not significantly reduced the viability til the E:T ratio 50:1; while total 4T1 cell’s viability reduced from E:T 10:1 and 4T1 non-CSC’s viability reduced from E:T 20:1.
At the E:T ratio of 20:1, viability was 65.87 ± 5.00% for total 4T1 cells, 56.23 ± 9.16% for CSC-like populations, and 59.42 ± 9.56% for non-CSCs. Notably, CSC-like populations showed significantly reduced viability compared to total 4T1 cells at several E:T ratios, while differences between CSC-like and non-CSC populations were less pronounced. These results suggest that γδT cells exert cytotoxic effects on both stem-like and non-stem tumor cell populations, with CSC-like populations displaying a tendency toward increased sensitivity under the tested conditions.
Cell cycle redistribution following γδT co-culture and drug treatment
Cell cycle distribution of 4T1 cells was evaluated by flow cytometry using the PI/RNase Staining Buffer Kit after co-culture with γδT cells (E:T = 20:1), in the presence or absence of APAP and MET (Figure 5). The percentages of cells in Sub-G1 (<G1), G1/G0, S, G2, and >G2 phases were analyzed to determine the effects of γδT-mediated cytotoxicity and drug treatment on cell cycle progression. Compared with the control group, co-culture with γδT alone resulted in a slight decrease in the Sub-G1 population (3.61 ± 0.29% vs. 5.26 ± 0.84%) and G1/G0 phase (59.63 ± 0.64% vs. 60.37 ± 0.81%), while the S phase was slightly increased (29.30 ± 0.20% vs. 28.33 ± 1.16%), suggesting limited effects on cell cycle arrest under this condition.
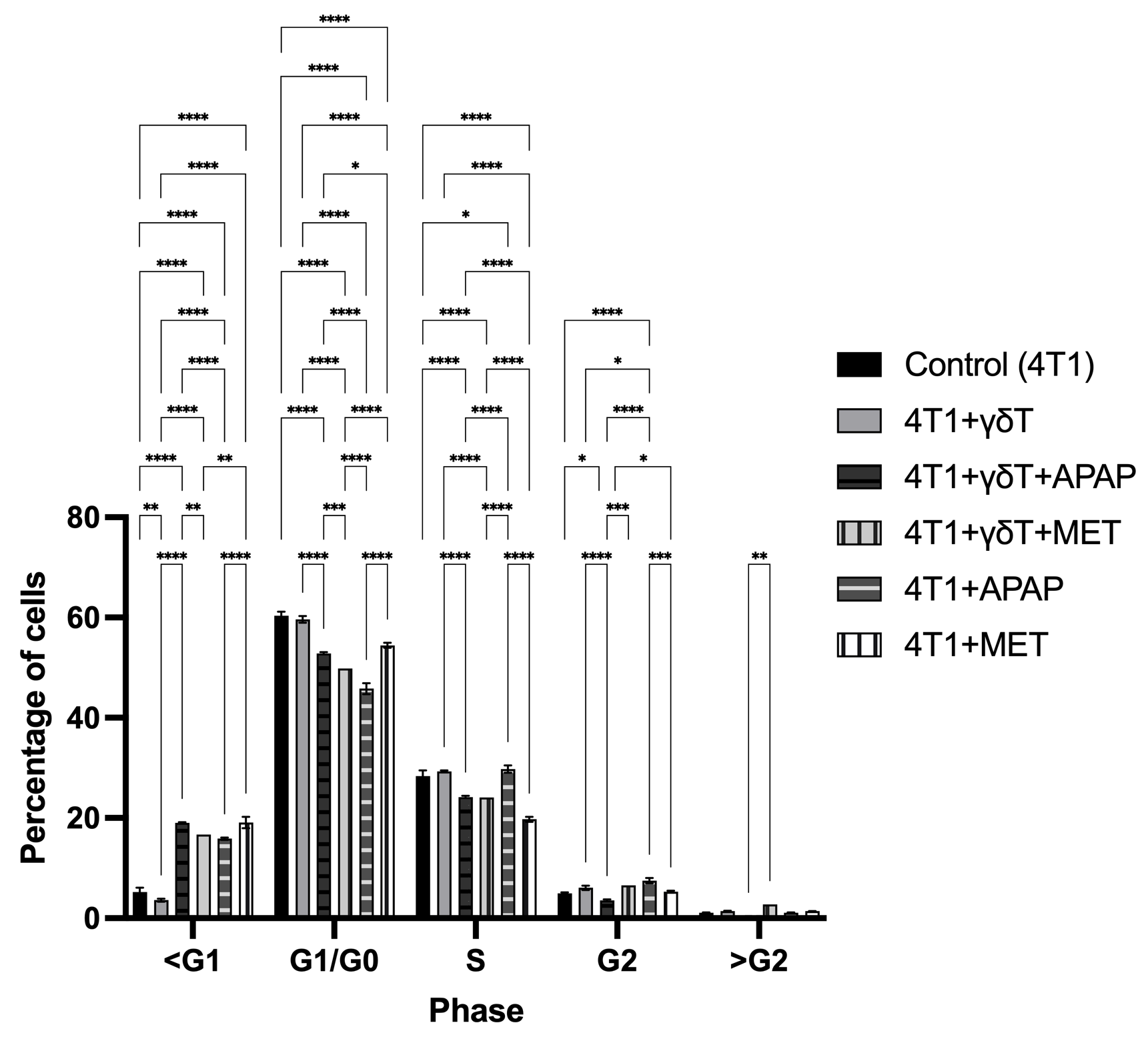
Cell cycle distribution of 4T1 cells after co-culture with γδT cells (E:T = 20:1) in the presence or absence of APAP and MET. Cell cycle progression was analyzed by flow cytometry using the PI/RNase Staining Buffer Kit, and the percentages of cells in Sub-G1 (<G1), G1/G0, S, G2, and >G2 phases were determined. Data are presented as mean ± SD (n = 3). Co-culture with γδT alone showed limited effects on cell cycle distribution compared to the control group. In contrast, APAP and MET treatments increased the Sub-G1 population and reduced the G1/G0 fraction, indicating enhanced apoptotic cell death rather than cell cycle arrest. APAP showed a stronger effect in promoting Sub-G1 accumulation under γδT co-culture conditions, while MET also maintained evident cytotoxicity in both single-treatment and co-culture settings. No marked differences were observed in the G2 and >G2 phases among the groups. Statistical significance between groups is indicated as p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
Notably, the addition of APAP markedly increased the Sub-G1 population in both the 4T1+γδT+APAP group (19.03 ± 0.15%) and the APAP-only group (15.87 ± 0.23%) compared to control and γδT alone, indicating enhanced cell death and apoptotic induction. This increase was accompanied by a substantial reduction in the G1/G0 population (52.83 ± 0.25% and 45.80 ± 1.08%, respectively) and a decrease in S phase cells, suggesting that APAP promoted apoptotic cell loss rather than cell cycle progression.
Similarly, treatment with MET alone also increased the Sub-G1 fraction (19.10 ± 1.13%) and reduced the G1/G0 population (54.45 ± 0.50%), indicating cytotoxic activity. However, the combination of γδT and MET showed a slightly lower Sub-G1 percentage (16.70%) compared to MET alone, while maintaining reduced G1/G0 levels (49.80%), suggesting that MET-induced cytotoxicity remained evident in the co-culture system.
No marked differences were observed in the G2 and >G2 populations among the groups, indicating that the major treatment-related changes occurred primarily in the Sub-G1 and G1/G0 phases.
Overall, these results suggest that APAP and MET both increase apoptotic cell death in 4T1 cells, as reflected by the expansion of the Sub-G1 population, while APAP appears to exert a stronger effect in the γδT co-culture condition.
Regulation of CSC-like population enrichment by γδT, APAP, and MET in 4T1 cells
The enrichment of CSC-like populations in 4T1 cells was evaluated by flow cytometry using CD44 and CD24 staining under co-culture conditions with γδT cells, in the presence or absence of APAP and MET (Figure 6). CSC-like populations were operationally defined as CD44CD24 cells based on surface-marker expression. Two analyses were performed: (A) the proportion of CSC-like populations within the total tumor cell population regardless of viability status, and (B) the proportion of CSC-like populations within surviving tumor cells (7-AAD-).
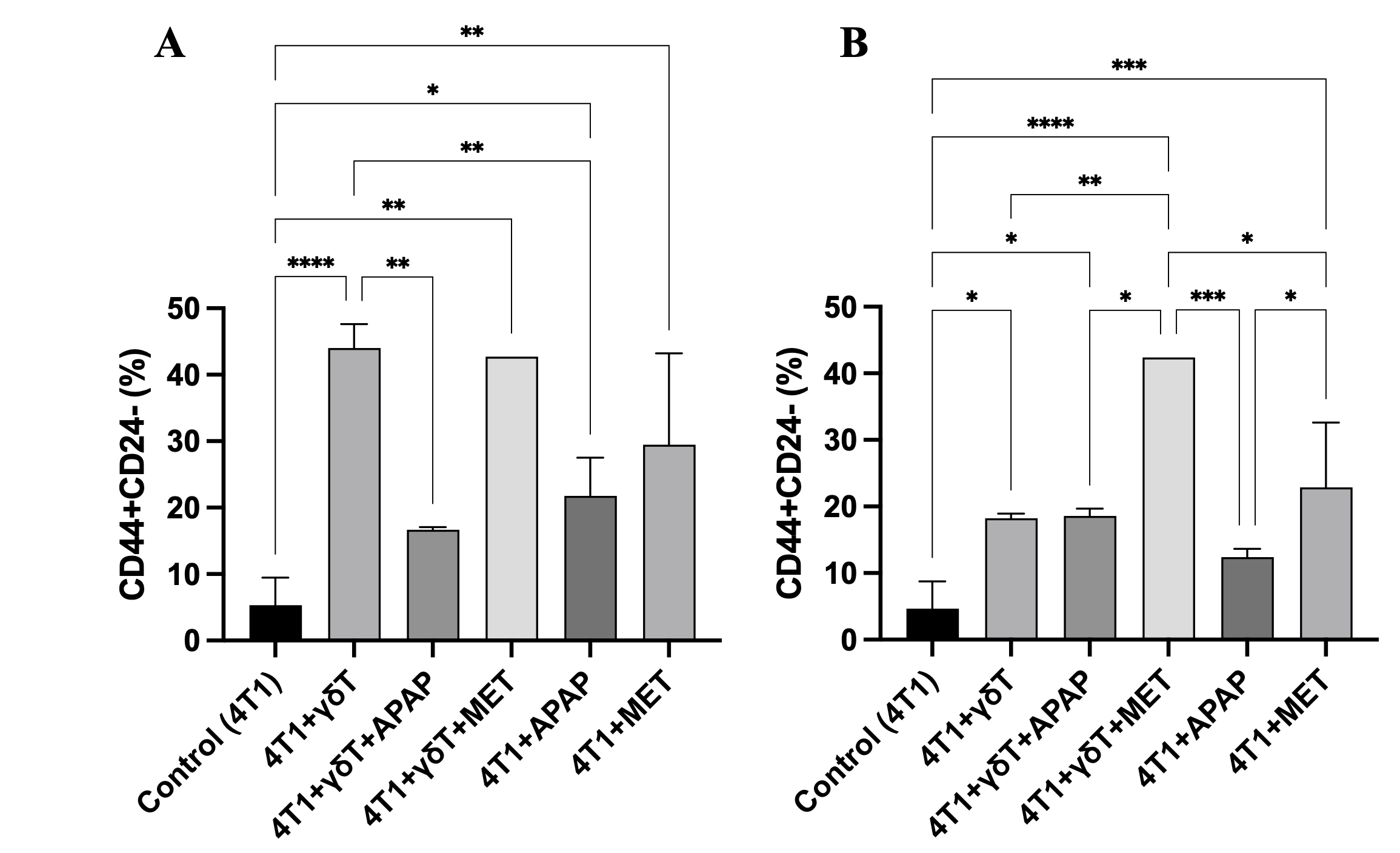
Enrichment of CSC-like populations (CD44+CD24−) in 4T1 cells after co-culture with γδT cells in the presence or absence of APAP and MET. CSC-like population proportions were evaluated by flow cytometry using CD44 and CD24 staining. (A) Percentage of CSC-like populations within the total 4T1 tumor cell population regardless of viability status. (B) Percentage of CSC-like populations within the surviving 4T1 cell population (7-AAD-). Data are presented as mean ± SD (n = 3–6 per group). Co-culture with γδT markedly increased CSC-like population enrichment compared to the control group. APAP treatment also increased the CSC-like fraction compared to control, although to a lesser extent than γδT. The addition of γδT to APAP-treated cells reduced CSC-like population accumulation, but this reduction was not statistically significant compared to APAP alone. In contrast, MET treatment maintained high CSC-like population proportions, particularly in the γδT+MET group, indicating persistent enrichment of CSC-like cells among surviving tumor cells. Statistical significance between groups is indicated as p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
In the total tumor cell population (Figure 6A), the control group showed a low baseline CSC-like population proportion (5.33 ± 4.13%). Co-culture with γδT markedly increased the CSC-like fraction to 44.04 ± 3.57%, indicating strong selective enrichment of the CD44CD24 population. APAP treatment increased the proportion of the CSC-like population (CD44CD24) in 4T1 cells. Although this increase was less pronounced than that induced by γδT-mediated selective pressure, it remained markedly higher than that of the control group. The APAP-only group showed a CSC-like population proportion of 21.79 ± 5.72%, which was substantially elevated compared to the baseline level in the control group (5.33 ± 4.13%, about 4 times more). The addition of γδT in the γδT +APAP group resulted in a lower CSC-like population proportion (16.68 ± 0.41%), suggesting a reduction in APAP-associated CSC-like population accumulation; however, this decrease was not statistically significant compared to the APAP-only group.
In contrast, MET treatment maintained a high CSC-like fraction. The γδT+MET group showed a CSC-like population proportion of 42.70%, which was comparable to γδT alone, indicating that MET did not reduce γδT-mediated CSC-like population enrichment. MET alone also exhibited a relatively high CSC-like population proportion (29.49 ± 13.75%), further suggesting that MET may favor the persistence of the CSC-like population.
A similar pattern was observed when only viable tumor cells (7-AAD-) were analyzed (Figure 6B). The baseline CSC-like population proportion in the control group was 4.67 ± 4.09%, which increased to 18.24 ± 0.68% after co-culture with γδT. In comparison with the corresponding results within the total 4T1 tumor cell population, the ratio of viable CSC-like populations in γδT group reduced remarkably (by half). The γδT+APAP group showed a comparable CSC-like population proportion (18.59 ± 1.11%) with the γδT group, and higher than APAP alone which was 12.39 ± 1.24%.
Notably, MET again showed the strongest association with CSC-like population enrichment in viable cells, with the γδT+MET group exhibiting the highest CSC-like population proportion (42.40%) and MET alone remaining markedly elevated (22.90 ± 9.74%). These results suggest that although MET enhanced apoptotic cell death in previous assays, the surviving tumor cells were substantially enriched in the CSC-like phenotype.
Overall, these findings demonstrate that γδT, APAP, MET treatments strongly enrich the CSC-like CD44CD24 population in 4T1 cells. APAP partially reduces this enrichment in the total tumor population but has limited effects among surviving cells, whereas MET is associated with persistent or enhanced CSC-like population accumulation, particularly in viable tumor cells.
Discussion
The present study differs conceptually from our previous work in human luminal breast cancer models because the highly aggressive 4T1 model represents a biologically distinct tumor system characterized by enhanced metastatic potential, increased stemness-associated properties, and greater adaptive responses to therapeutic stress. Therefore, the current investigation was designed not merely to examine drug-induced changes in γδT-mediated cytotoxicity, but also to evaluate how immune-selective pressure reshapes tumor-cell composition and influences residual CSC-like populations in a more aggressive breast cancer setting. These biological characteristics address questions distinct from those explored in luminal breast cancer models.
An important consideration in the present study is the use of human-derived γδT cells in co-culture with murine 4T1 breast cancer cells. Unlike conventional αβ T cells, γδT cells recognize transformed cells predominantly through MHC-independent mechanisms involving stress-induced ligands, NKG2D signaling, death receptor pathways, and perforin/granzyme-mediated cytotoxicity rather than classical peptide–MHC interactions11,26. These characteristics provide a biological basis for exploratory xenogeneic tumor-killing assays. Although direct evidence for human γδT–murine tumor interactions remains limited, studies involving other innate cytotoxic lymphocytes, such as NK cells, have demonstrated antitumor activity across species through conserved MHC-unrestricted pathways27,28. Nevertheless, the present co-culture system should be interpreted primarily as an exploratory model for assessing tumor-cell responses under immune-selective pressure rather than a direct representation of physiological human–mouse immune interactions.
In the present study, we demonstrated that APAP and MET distinctly modulate γδT cell (γδT)-mediated antitumor activity against 4T1 murine breast cancer cells. While expanded primary γδT cells exerted strong cytotoxicity against 4T1 cells, APAP was associated with reduced γδT-mediated tumor killing, whereas MET displayed a more complex dual role characterized by potent direct cytotoxicity but simultaneous enrichment of surviving CSC-like populations. However, because γδT-cell functional assays were not performed, it remains unclear whether these effects resulted primarily from altered effector-cell function, tumor susceptibility, or both. These findings highlight that commonly used metabolic-modulating agents can substantially reshape tumor–immune interactions and may critically influence the therapeutic outcome of γδT cell-based immunotherapy4,29,30.
Consistent with previous studies, γδT cells showed robust antitumor activity against breast cancer cells, reflected by increased PI-positive populations, elevated apoptotic fractions, and reduced cell viability in co-culture conditions. γδT cells recognize malignant cells in an MHC-independent manner and mediate cytotoxicity through perforin/granzyme release, Fas/FasL interactions, TRAIL signaling, and secretion of pro-inflammatory cytokines such as IFN-γ and TNF-α4,29. Recent evidence has further emphasized the therapeutic relevance of γδT cells in triple-negative breast cancer (TNBC), where they can target both bulk tumor cells and stem-like subpopulations while bypassing conventional antigen-presentation constraints4,11. However, the function of γδT cells in breast cancer is context-dependent, and both anti-tumor and pro-tumor roles have been described depending on the tumor microenvironment and immune composition29,31.
Interestingly, our results showed that the CD44CD24 CSC-like population displayed greater sensitivity to γδT-mediated cytotoxicity than the total 4T1 population, which contrasts with the classical concept that cancer stem cells are broadly resistant to therapy. This observation is nevertheless supported by recent reports demonstrating that γδT cells can preferentially recognize and eliminate breast CSCs through upregulation of MHC-I and CD54/ICAM-1 expression, stress-induced ligands, including NKG2D ligands, phosphoantigen-associated pathways, and altered death receptor signaling11,12. In aggressive breast tumors, CSCs often maintain elevated metabolic stress signatures that may increase their susceptibility to immune-mediated killing despite resistance to chemotherapy or radiotherapy. Therefore, immune sensitivity and conventional therapeutic resistance should be considered as biologically distinct properties rather than mutually exclusive phenotypes11,12.
One of the most notable findings of this study is that APAP was associated with reduced tumor killing under γδT co-culture conditions, despite inducing moderate apoptosis when used alone. APAP significantly reduced the proportion of PI-positive cells in the γδT co-culture system and partially decreased γδT-associated CSC-like population enrichment in the total tumor population. However, Annexin V/PI analysis and cell cycle profiling showed increased early apoptosis and marked Sub-G1 accumulation in APAP-treated groups. This apparent discrepancy may reflect the distinction between apoptotic initiation and effective immune-mediated terminal elimination. APAP is known to induce oxidative stress through glutathione depletion, mitochondrial protein adduct formation, and subsequent mitochondrial dysfunction, which may sensitize tumor cells to apoptotic signaling21. However, excessive oxidative stress may also impair apoptosis execution, immune synapse formation, and death receptor signaling, thereby reducing the efficiency of γδT-mediated killing32,33,34. In addition, the APAP-induced increase in CSC-like populations suggests that sublethal pharmacological stress may selectively preserve more stem-like and immunologically adaptable tumor cells. Given that APAP is widely administered as supportive care during cancer treatment, its potential to alter antitumor immune responses deserves greater clinical attention.
MET showed a substantially different profile. As a single agent, MET induced strong apoptosis, markedly increased PI positivity, and caused severe disruption of 4T1 cell morphology, supporting its well-established role as a metabolic stressor. MET inhibits mitochondrial complex I, disrupts oxidative phosphorylation, activates AMPK signaling, and suppresses anabolic pathways required for tumor growth17,18. In our co-culture model, MET further enhanced early apoptosis and induced extensive tumor cell detachment when combined with γδT, suggesting stronger overall cytotoxic stress. However, MET was also associated with the highest enrichment of surviving CD44CD24 CSC-like populations, particularly in the γδT+MET group. This paradox indicates that while MET efficiently eliminates a large proportion of tumor cells, the residual population may become highly enriched for metabolically adaptable CSC-like populations. Similar findings have been reported in breast cancer models where metabolic therapies selectively eliminate differentiated tumor cells but allow stem-like subsets to survive and dominate the residual tumor compartment35,36,37,38. Thus, MET may improve short-term tumor killing while simultaneously increasing the long-term risk of persistence or relapse if CSC-targeting strategies are not incorporated.
The cell cycle analysis further supports this interpretation. γδT alone exerted limited effects on classical cell cycle arrest, whereas both APAP and MET significantly increased the Sub-G1 population and reduced the G1/G0 fraction, indicating apoptosis-driven cell loss rather than checkpoint-mediated growth arrest. The absence of major changes in G2 or >G2 phases suggests that cytotoxic elimination rather than proliferative arrest is the dominant mechanism in this system. Notably, the discrepancy between Annexin V/PI apoptosis assays, PI-based viability measurements, and representative bright-field microscopic images in the γδT+MET group highlights an important technical issue: extensive cell detachment during strong cytotoxic responses may lead to underestimation of total cell death when only adherent or retained cells are analyzed. This is particularly relevant in co-culture models involving highly cytotoxic immune effectors and should be carefully considered when interpreting flow cytometric viability data.
Taken together, our findings indicate that γδT cell therapy alone exerts strong antitumor activity against murine breast cancer cells, but its efficacy can be significantly modulated by commonly used pharmacological agents. APAP may weaken effective immune-mediated tumor elimination despite promoting apoptotic signaling, whereas MET enhances cytotoxic stress but simultaneously selects for persistent CSC-like survivors with potentially greater therapeutic resistance. These results emphasize that evaluating direct tumor cell death alone is insufficient; the composition and biological properties of residual surviving cells must also be considered when designing combinational immunotherapy strategies. From a translational perspective, the routine use of supportive medications such as APAP during cancer treatment should be more carefully evaluated, and metabolic agents such as MET may require rational combination with CSC-targeting approaches or immune escape blockade to maximize durable therapeutic benefit.
Besides, several limitations should be acknowledged. First, the use of human-derived γδT cells against murine 4T1 cells represents an inherent limitation of the present study. Although γδT-mediated cytotoxicity relies predominantly on conserved innate-like recognition mechanisms rather than classical MHC restriction, species-dependent differences in ligand expression and signaling pathways may influence effector–target interactions11,26. Therefore, future studies using syngeneic murine γδT systems will be necessary to further validate the translational relevance of the current findings. Second, CSC-related observations in this study were based exclusively on CD44CD24 surface-marker expression. Although this phenotype is commonly associated with stemness-related characteristics in breast cancer, surface-marker analysis alone cannot establish functional stemness properties. Additional functional assays such as mammosphere formation, ALDH activity, limiting-dilution tumorigenicity, or serial transplantation would be required to confirm true stem-cell behavior. Third, the pharmacological concentrations used in this study exceeded clinically achievable plasma concentrations of APAP and MET. While these concentrations were intentionally selected to establish robust metabolic stress conditions and permit measurable biological effects within the in vitro co-culture system, caution should be exercised when extrapolating these findings directly to clinical settings. Future studies should incorporate dose–response analyses across physiologically relevant concentration ranges and pharmacokinetically informed experimental designs. Next, the precise mechanisms underlying APAP- and MET-associated modulation of tumor-cell killing were not directly investigated. In particular, functional characteristics of γδT cells, including degranulation capacity (CD107a), cytokine secretion (IFN-γ and TNF-α), perforin/granzyme expression, and activation or exhaustion phenotypes, were not assessed. In addition, flow cytometric viability analyses were performed only on adherent tumor cells, while floating cells were not collected. Since highly damaged or dead cells may preferentially detach following strong cytotoxic treatment, the measured viability may underestimate the overall extent of tumor cell killing. Future studies should include both adherent and floating populations and incorporate tumor-specific labeling markers to accurately distinguish tumor cells from effector cells during co-culture experiments.
Conclusion
APAP and MET differentially modulate γδT cell-mediated antitumor activity against 4T1 breast cancer cells. While APAP was associated with reduced γδT-mediated tumor killing despite promoting apoptotic signaling, MET induced strong direct cytotoxicity but simultaneously enriched surviving CSC-like populations, particularly among viable residual cells. These findings suggest that therapeutic evaluation should consider not only immediate tumor cell killing but also adaptive changes in residual tumor-cell composition and CSC-like enrichment following immune-selective pressure. Optimizing γδT cell-based immunotherapy may therefore require combination strategies targeting both CSC-like population persistence and immune escape mechanisms.
Abbreviations
7-AAD: 7-Aminoactinomycin D; ANOVA: Analysis of Variance; APAP: Acetaminophen (Paracetamol); CSC-like: Cancer Stem Cell-like population; DMSO: Dimethyl Sulfoxide; E:T: Effector-to-Target Ratio; FACS: Fluorescence-Activated Cell Sorting; FBS: Fetal Bovine Serum; FCM: Flow Cytometry; IACUC: Institutional Animal Care and Use Committee; IC50: Half-Maximal Inhibitory Concentration; MET: Metformin; mRNA: Messenger Ribonucleic Acid; PBMCs: Peripheral Blood Mononuclear Cells; PBS: Phosphate-Buffered Saline; PI: Propidium Iodide; RT-qPCR: Reverse Transcription Quantitative Polymerase Chain Reaction; SD: Standard Deviation; SPF: Specific Pathogen-Free; TNBC: Triple-Negative Breast Cancer; γδT: Gamma delta T cells .
Acknowledgments
None.
Author’s contributions
N.C.T. conceived the study, performed or participated in all experiments, analyzed the data, and drafted the manuscript. N.T.H. managed the cell culture and treatment. P.V.P. revised the manuscript and provided conceptual guidance. All authors read and approved the final manuscript.
Funding
This research is funded by Vietnam National University, Ho Chi Minh City (VNU-HCM) under grant number CB2025-18-26: “Evaluation of the effect of Paracetamol on the immune escape of breast cancer cells”, Project leader: MSc. Truong Chau Nhat within the framework of the Program titled "Strengthening the capacity for education and basic scientific research integrated with strategic technologies at VNU-HCM, aiming to achieve advanced standards comparable to regional and global levels during the 2025-2030 period, with a vision toward 2045. This research is also funded by VNUHCM-US Stem Cell Institute, University of Science, VNU-HCM, Viet Nam under grant number NCKH-SCI.09/25”.
Availability of data and materials
Data and materials used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
Peripheral venous blood (15 mL) was collected from healthy adult donors. The collection protocol was strictly conducted in adherence to the Declaration of Helsinki and was approved by the Institutional Ethics Committee (NCKH-SCI.09/25). Written informed consent was obtained from all participants prior to inclusion.
Consent for publication
Not applicable.
Declaration of generative AI and AI-assisted technologies in the manuscript preparation process
During the preparation of this work the author(s) used Gemini in order to improve the language and readability of their paper. After using this tool/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the published article.
Competing interests
The authors declare that they have no competing interests.